Important Discoveries of the neurohypophyseal system
- 1794 J. P. Frank distinguished diabetes insipidus from diabetes mellitus.
- 1895 Olliver and Schafer produced an extract from the pituitary which caused a rise in blood pressure.
- 1910 A. Frank associated diabetes insipidus with a hypofunction of the posterior pituitary.
- 1913 Camus and Roussy produced diabetes insipidus in dogs by hypothalamic lesions, without injury to the pituitary.
- 1913 Farmi and Von Den Velden established the therapeutic effect of extracts of the posterior pituitary in diabetes insipidus.
- 1948 Verney showed that the activity of the hypothalamic-neurohypophyseal system was regulated by the osmotic concentration of the blood in the hypothalamus.
- 1953/55 Du Vigneaud isolated and synthesized vasopressin and oxytocin.
What is the hypothalamus
The hypothalamus has a diameter of only 2.5 cm and makes up 1/300th of the weight of the brain. Nevertheless, this tiny organ coordinates and regulates most of the vital processes of life. Modulations are effected through neural and neuro-endocrine routes.
The hypothalamus controls the endocrine system primarily through neurosecretions acting on the anterior pituitary. These are usually of a stimulatory nature (releasing factors), but may occasionally be of an inhibitory nature (inhibitory factors).
Embryology and Gross Anatomy of the Hypothalamus
In the human embryo, the anlagen of the forebrain (prosencephalon), midbrain (mesencephalon) and hindbrain (rhombencephalon) become distinguishable even before closure of the rostral end of the neural groove.
When the brain becomes a closed tube (neural tube), these three subdivisions develop into the primary brain vesicles. Early in the fourth week (when the length of the embryo is approximately 3 mm) a dorsal groove forms in the prosencephalic vesicle
subdividing it into the telencephalon, with the primitive cerebral hemispheres, and the diencephalon, with the anlage of the eyes, i.e., the optic vesicles.
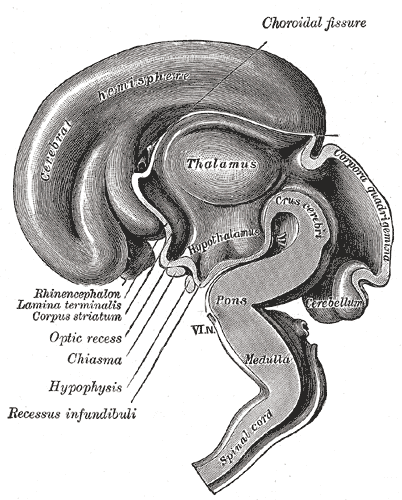
In embryos 7-8 mm in length, the anlagen of the pineal gland and the neurohypophysis appear as small outgrowths of the roof and the floor of the diencephalon, respectively. The Diencephalon can be divided into four more or less distinct major parts.
The dorsally situated epithalamus (1) comprises the epithelial roof of the third ventricle, the striae medullares thalami, the habenular trigones and the pineal gland. The thalamus (2) extends basally to the sulcus hypothalamicus Monroi (sulcus diencephalicus ventralis).
Its medial and lateral portions are separated from each other by the internal medullary lamina. Both contain a number of morphologically distinct nuclear masses. The subthalamus (3) or ventral thalamus is bounded dorsally by the thalamus, ventrally and laterally by the pars peduncularis of the internal capsule, and medially and rostrally by the hypothalamus.
It includes the subthalamic nucleus, the zona incerta, the nucleus of the tegmental field of Forel and, caudally, the rostral parts of both the substantia nigra and the red nucleus.
The subthalamus is continuous with the tegmentum and the basilar portions of the mesencephalon and must therefore be considered as part of the extrapyramidal motor system. Because of its abundance of myelinated nerve fibers, the subthalamus can easily be distinguished from the hypothalamus (4) which contains mainly nonmyelinated nerve fibers and a much richer vascular network. The mamillary bodies, although usually considered as part of the hypothalamus, belong to the subthalamus. They receive myelinated fibers from the fornix. The Hypothalamus comprises the lower parts of the ventral walls of the third ventricle as well as the structures of the ventricular floor (chiasma opticum, tuber cinereum and infundibulum). It is separated from the pars telencephalica of the third ventricle by a line connecting the optic chiasm with the posterior edge of the anterior commissure.
Caudal and lateral to the hypothalamus is the subthalamus. An inferior view of the brain shows that the hypothalamus is bounded anteriorly by the optic chiasma; laterally by the optic tracts and posteriorly by the mamillary bodies.
The hypothalamus consists of a diffuse matrix of rather small cells which form the central gray substance. Within this matrix, a number of more or less well distinguishable nuclear masses (= nuclei) permit the hypothalamus to be divided morphologically into three regions: The supraoptic region (1) lies directly above the optic chiasma. It contains the supraoptic and paraventricular nuclei which are bridged by small groups of cells.
Both these nuclei consist of large, often bipolar and darkstaining neurons which show cytoplasmic inclusions of colloidal material and peripherally distributed Nissl substance. The majority of their efferent nerve fibers make up the supraoptico-hypophyseal tract, which extends to the posterior lobe of the hypophysis. A minority of the axons, however, terminate within the nuclei themselves or within the infundibulum.
The central gray substance of the tuberal region (2) contains the nucleus tuberis infundibularis, the nucleus hypothalamicus ventromedialis and the so-called periventricular area.
These nuclear masses consist of small, ovoid cells intermingled with some larger elements. They are the site of origin of the tubero-hypophyseal tract, most of whose nerve fibers terminate in close proximity to the specialized blood vessels of the hypophyseal portal system.
This in turn extends to the sinusoidal capillary network of the anterior lobe of the hypophysis. Finally, a lateral area (3) of the tuber cinereum contains the tubero-mamillary nucleus and the lateral tuberal nuclei. Little is known about their nerve fiber connections.
The Blood Supply of the hypothalamus is ensured mainly by the anterior cerebral artery and the posterior communicating artery. Small branches extend from these vessels to form an intricate network within the nervous parenchyma. The supraoptic and paraventricular areas are characterized by a particularly dense capillary network which is supplied by branches of the anterior cerebral artery, more specifically the supraoptic-paraventricular arteries. The supraoptic nucleus also receives small branches from the superior hypophyseal artery. The nuclear masses of the tuber cinereum share the general, less dense vascular network of the hypothalamus, which is supplied partly by the anterior cerebral arteries and partly by the posterior communicating arteries.
The blood supply of this region is therefore ensured by at least two large pairs of arteries, which also supply the mammillary bodies. The hypophysis (pituitary gland) itself receives its blood supply from the superior and inferior hypophyseal arteries, which are branches of the internal carotid. There are no direct connections between the vascular networks of the various hypothalamic nuclei and the hypophysis.
The Neurohypophysis
The hypophysis has two major subdivisions which differ fundamentally in embryology, morphology, and function. The adenohypophysis (anterior pituitary gland) is derived from a dorsal outpocketing of the ectodermal buccopharyngeal region. Indeed, it demonstrates all the characteristics of an endocrine gland.
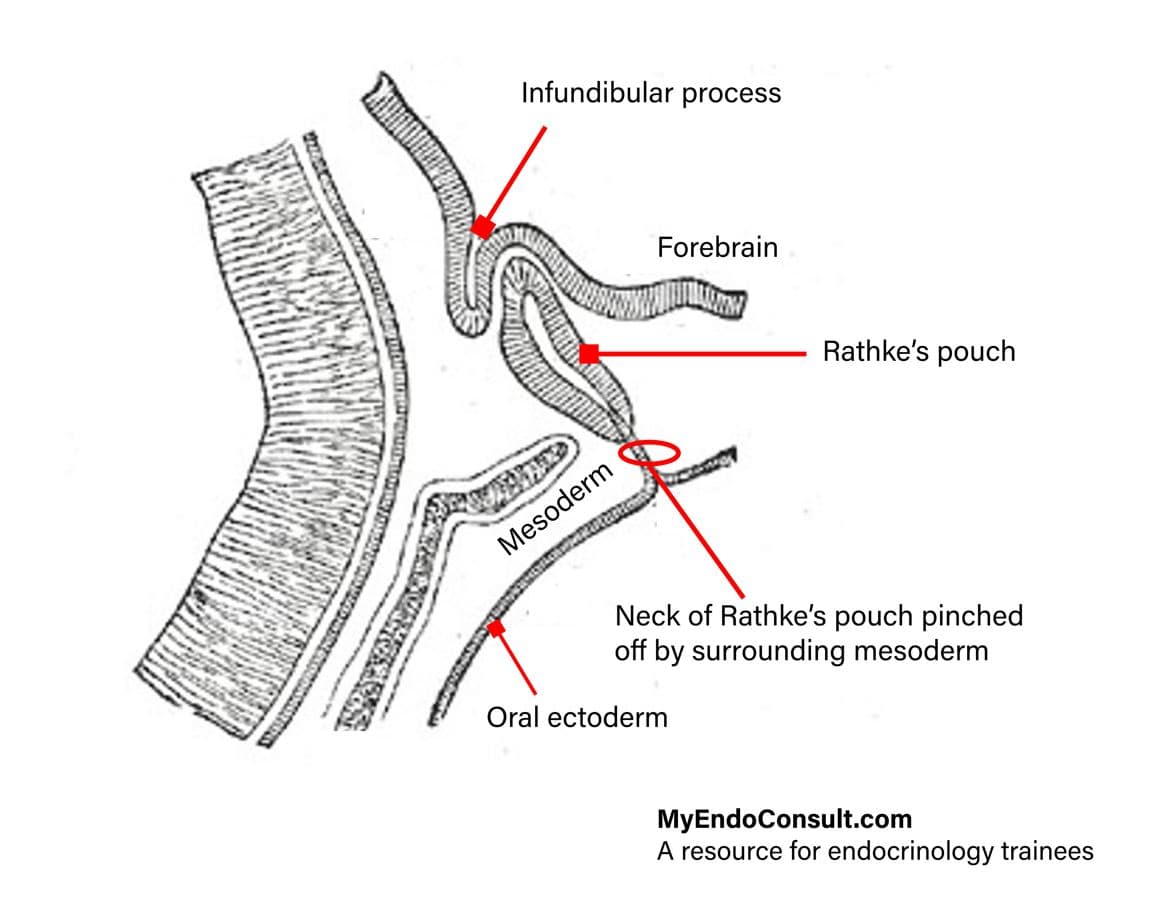
The neurohypophysis originates from the diencephalon. In the 7-8 mm-Iong human embryo, the neurohypophyseal anlage appears first as a thickening of the diencephalic floor. From this, a compact epithelial process grows downward behind the anlage of the adenohypophysis and develops into infundibulum and neural lobe.
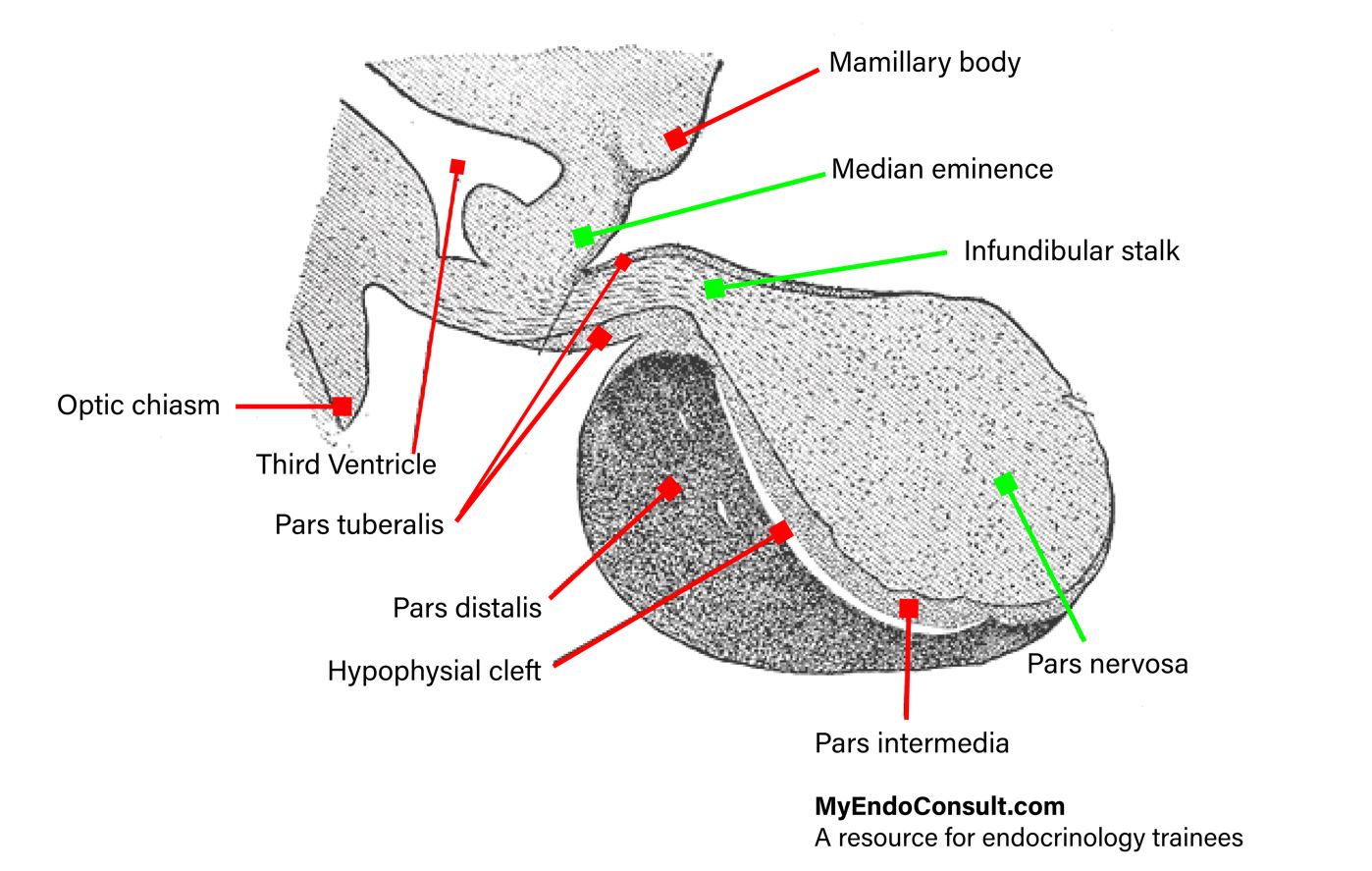
Gross Anatomy : The fully developed neurohypophysis has three subdivisions:
- The infundibulum (neural stalk, pars proximalis of the neurohypophysis) extends from the floor of the diencephalon down to the diaphragma sellae of the sphenoid bone. It forms the neurohypophyseal portion of the hypophyseal stalk. Its boundary towards the tuber cinereum is given by the hypothalamo-hypophyseal sulcus (sulcus tuberoinfundibularis). At the root of the infundibulum, the third ventricle evaginates to form the infundibular recess. The median eminence, a funnel-shaped extension of the tuber cinereum, must morphologically and functionally be considered as part of the infundibulum.
- The infundibular stalk is the border area between infundibulum and neural (posterior) lobe. It contacts with both the pars tuberalis and the pars intermedia of the adenohypophysis.
- The infundibular process (neural lobe, pars distalis of the neurohypophysis) is lodged within the sella turcica of the sphenoid bone, adjacent to the pars intermedia of the adenohypophysis.
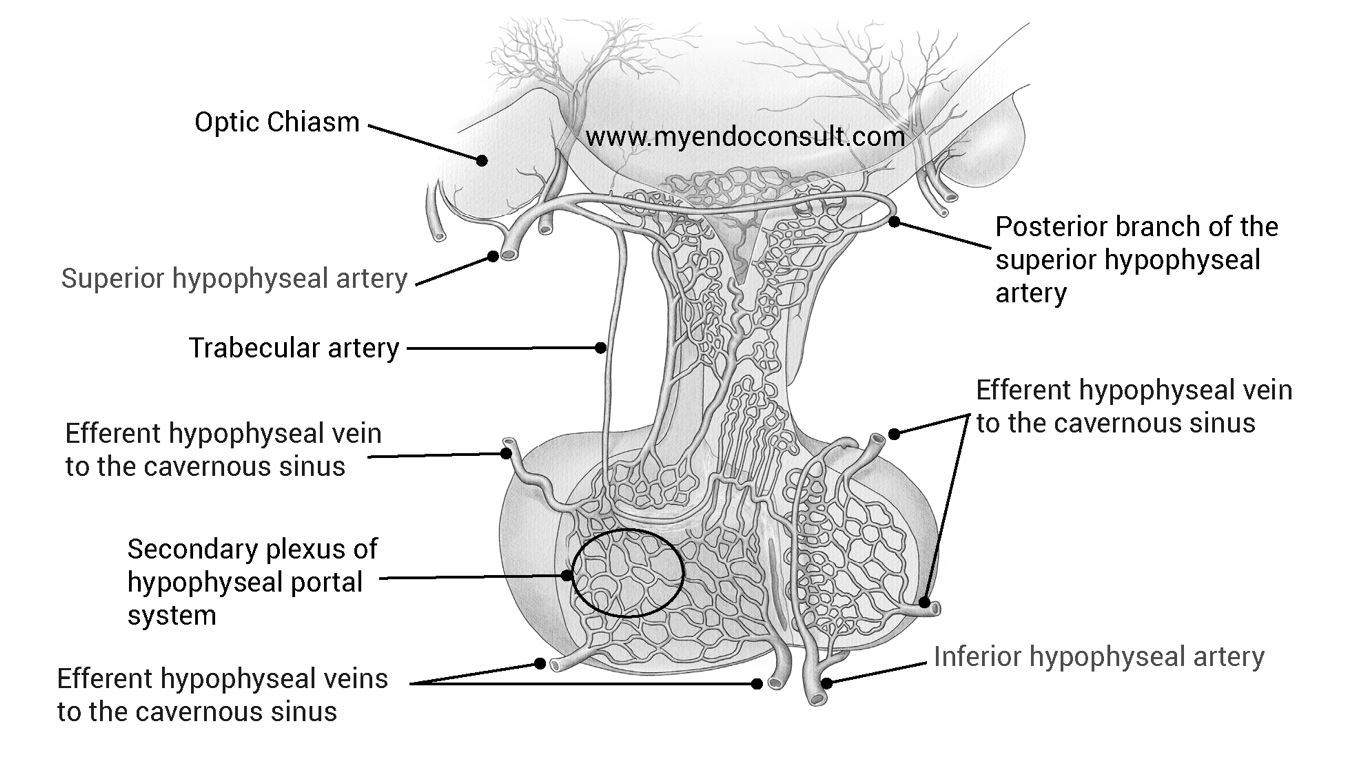
Blood Supply : The hypophysis is supplied by the left and right superior hypophyseal arteries which arise from the internal carotids near the origin of the ophthalmic arteries. Each superior hypophyseal artery divides into an anterior and a posterior branch (anterior and posterior superior hypophyseal artery, respectively). In the median eminence, the anterior branches form characteristic capillary loops, which drain into the hypophyseal portal vessels.
These in turn, supply the sinusoidal capillary network in the adenohypophysis. The anterior branches also supply the upper parts of the pars infundibularis of the adenohypophysis and, in addition, give rise to a small trabecular artery which passes directly into the anterior lobe where it anastomoses with the inferior hypophyseal artery.
The posterior branches break up into a capillary network within the infundibulum. The region of the tuber cinereum receives additional blood from branches of the posterior communicating arteries. The neural lobe and the lower portions of both the infundibulum and the pars infundibularis of the adenohypophysis are supplied by the inferior hypophyseal arteries. These are also branches of the internal carotids and anastomose with the trabecular arteries.
The venous blood is conveyed by capsular veins of the hypophysis into the anterior and posterior intercavernous sinuses which drain into the cavernous sinus. Thus, the hypophysis is surrounded by large blood channels anteriorly, posteriorly, and bilaterally (circular sinus).
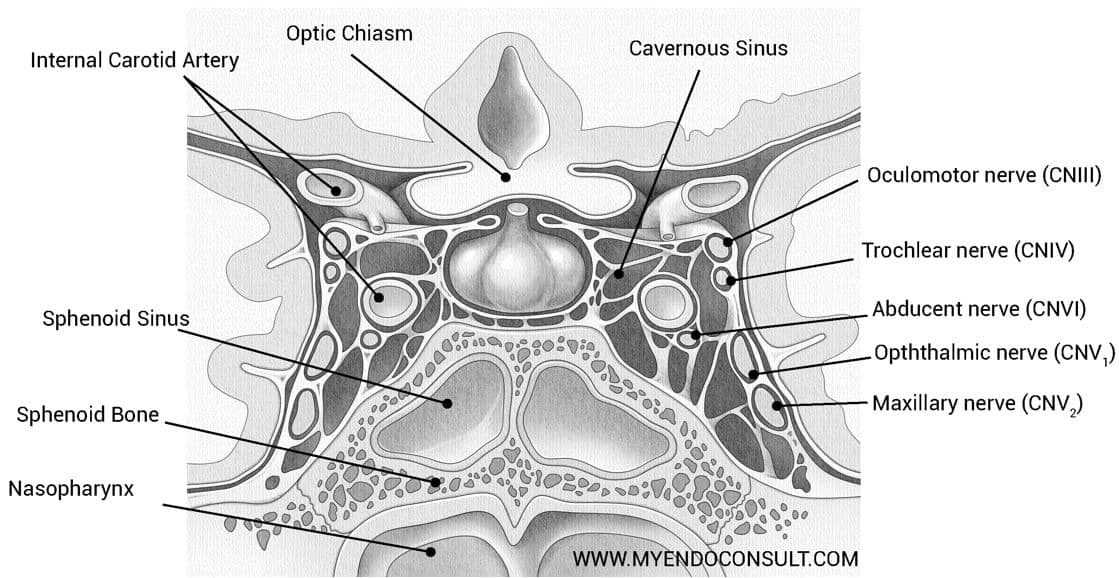
Intra- and suprasellar portions of the hypophysis are in close proximity to vital brain structures and blood vessels. Through the foramen hypophyseale of the diaphragma sellae, the sella turcica of the sphenoid bone opens into the middle cranial fossa. The lateral wall of the dura mater lining the sella is also the middle wall of the cavernous sinus. The hypophyseal stalk, which connects the intrasellar portions of the hypophysis with the hypothalamus, is situated immediately beneath the optic chiasma.
Although morphological variations of the chiasma are frequent, its close spatial relationship to the hypophyseal stalk is a constant feature. Tumors of the adenohypophysis can damage the decussating nerve fibers in the chiasma and lead to bitemporal hemianopsia. The proximal part of the infundibulum of the neurohypophysis encloses the infundibular recess of the third ventricle and thus gains close connections with the ventricular system of the brain.
In the region of the periinfundibular cistern, the hypophyseal stalk is also surrounded by cerebrospinal fluid. In addition, the infundibulum lies in the center of the arterial circle of Willis and in close proximity o the small but numerous arteries of the olfactory area. This area joins the infundibulum rostrally and laterally. Finally, the tuber cinereum and the two mamillary bodies are also situated close to the infundibulum.
Histology : In routine histological sections, the neurohypophysis appears as a mass composed of innumerable ill-defined fibers, among which a rather heterogeneous population of cells is distributed.
In sections stained with silver salts, a fine network of argyrophilic fibers is found to surround the blood vessels branching within the organ. Like the adenohypophysis, the neurohypophysis is enclosed by a relatively thick connective tissue capsule.
From this capsule, a thin septum rises between the anterior and posterior lobe (pars intermedia) towards the brain. This boundary may in parts, be erased by a penetration of basophilic cells from the pars intermedia of the adenohypophysis into the posterior lobe.
The neurohypophysis contains two major elements. The bulk of its substance is made up of about 100,000 unmyelinated nerve fibers, whose cell bodies are located in the hypothalamus. Distributed among these fibers are the pituicytes, which vary greatly in shape and size. Due to their structural relation to the axons, these cells must be considered as a special form of neuroglia. They commonly contain various amounts of pigment granules.
A broad perivascular space bounded on all sides by a basal lamina separates the nerve fiber endings and the pituicyte processes from the network of blood capillaries with fenestrated endothelia.
Light microscopy reveals the characteristic strongly positive Periodic Acid Schiff (PAS-reaction) of this perivascular area, which indicates the presence of large amounts of mucopolysaccharides.
In the electron microscope, finger-shaped ramifications from this space can often be traced for a good distance between the axons and the pituicytes.
The Hypothalamo-Neurohypophyseal Connections
The axons comprising the major part of the neurohypophyseal mass collectively form the hypothalamo-neurohypophyseal tract. Its cell bodies (perikarya) are located mainly within the supraoptic and paraventricular nuclei, whereas other hypothalamic nuclear masses apparently contribute only a few nerve fibers.
The axons pass via the infundibulum and the infundibular stalk to reach the posterior lobe where they terminate in intimate contact with the capillaries. When special staining methods are used, variously sized, intensively staining masses can be recognized in both the cell bodies of the hypothalamic nuclei and in their axons. These are the so called Herring bodies. In the electron microscope, these bodies have been found to be locally dilated portions of the axons where granules are densely packed with mitochondria and other cell organelles.
The granules are thought to contain the hormones synthesized on the granular endoplasmic reticulum (Nissl substance) and bound to a carrier protein in the Golgi complex. They are carried along the entire length of the axons into the posterior lobe. In addition to these neurosecretory granules with a diameter of about 100 to 200 nm, the nerve fiber endings also contain smaller vesicles of about 40 nm diameter. Such vesicles are normally found in synapses elsewhere in the nervous system. Their function at this site is still a matter of dispute.
The two hormones formed in the supraoptic and paraventricular nuclei are oxytocin and vasopressin. Since no neurosecretory granules have been demonstrated within the extensive perivascular space separating the capillary endothelia from the pituicytes and the nerve fiber endings, it must be assumed that the granules are dissolved while still within the axon terminals and that their content is emptied in molecular form into the cytoplasmic ground substance.
From there, after passing through the axonal cell membrane, the hormones reach the perivascular space rich in mucopolysaccharides and mucoproteins.
The function of the pituicytes in this process has not yet been clarified. Increasing evidence indicates, however, that these cells do not only have a supportive function but that they are largely involved in the metabolism of the neurosecretory cells and their axons.
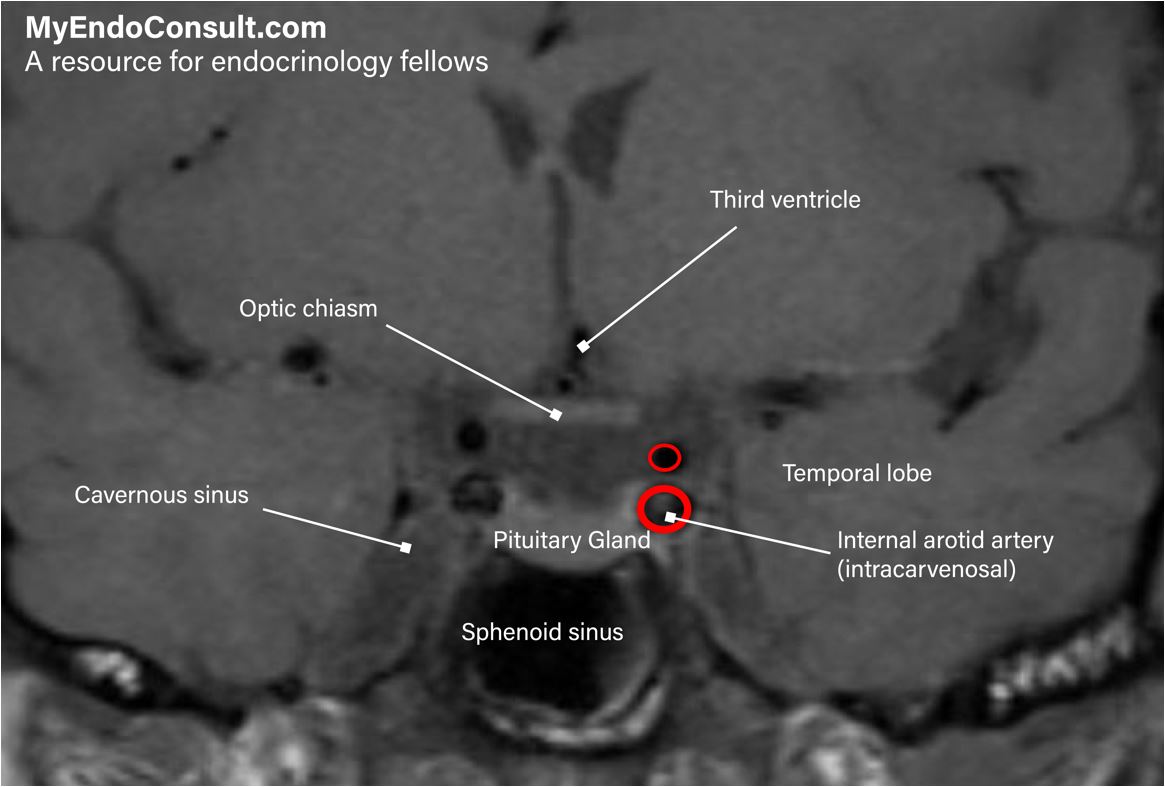
Practical Tip (Interpretation of pituitary imaging)
A simple guide for interpreting normal and abnormal findings in imaging of the hypothalamus and pituitary gland.